
Systematic Determination of Herpesvirus in Free-Ranging Cetaceans Stranded in the Western Mediterranean: Tissue Tropism and Associated Lesions

 , , , , , , , and
, , , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Herpesvirus Detection
2.3. Phylogenetic Analysis
2.4. Herpesvirus RNA Detection to Assess Active Replication
2.5. Herpesvirus and Morbillivirus Coinfection
2.6. Histopathological Study
3. Results
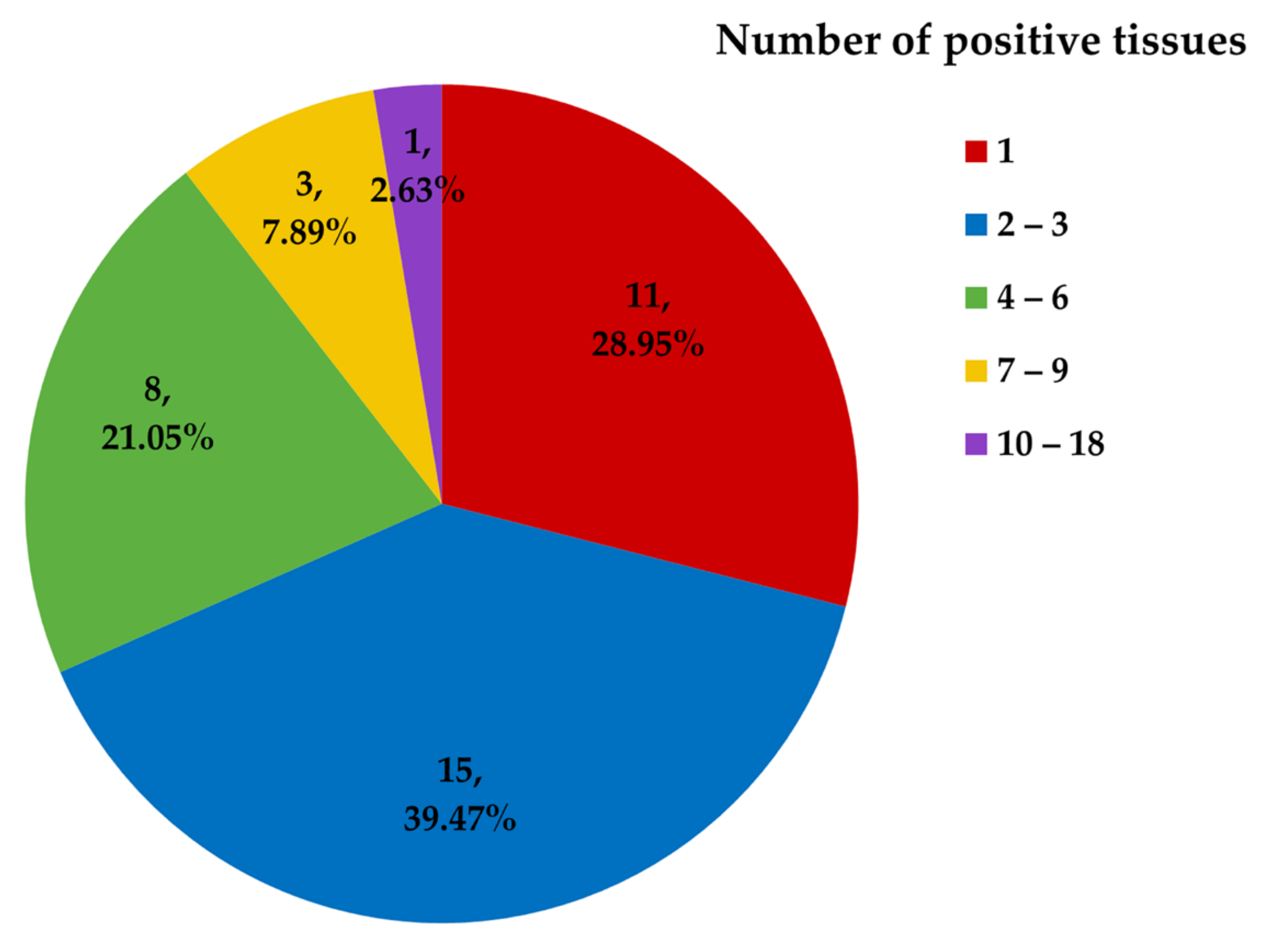
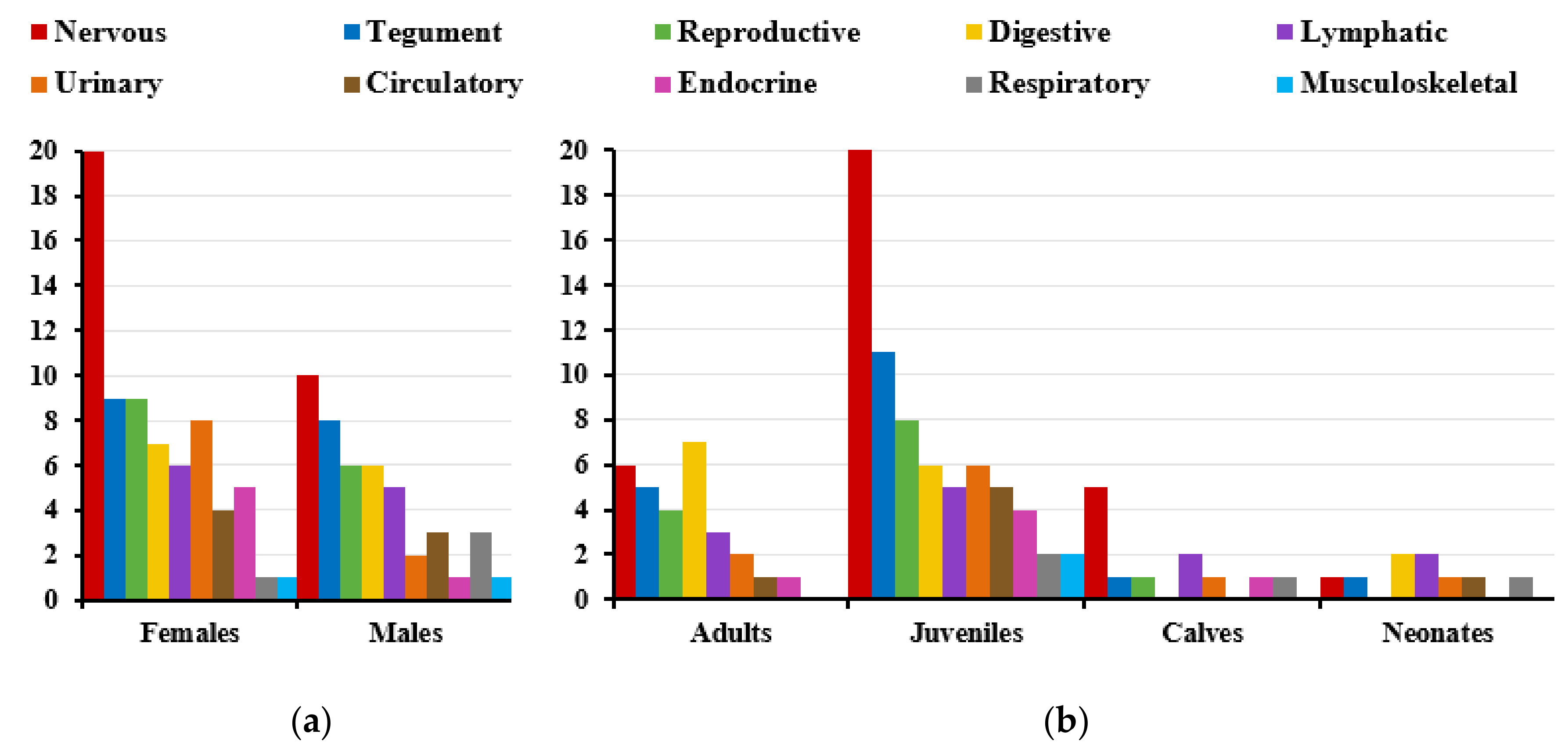
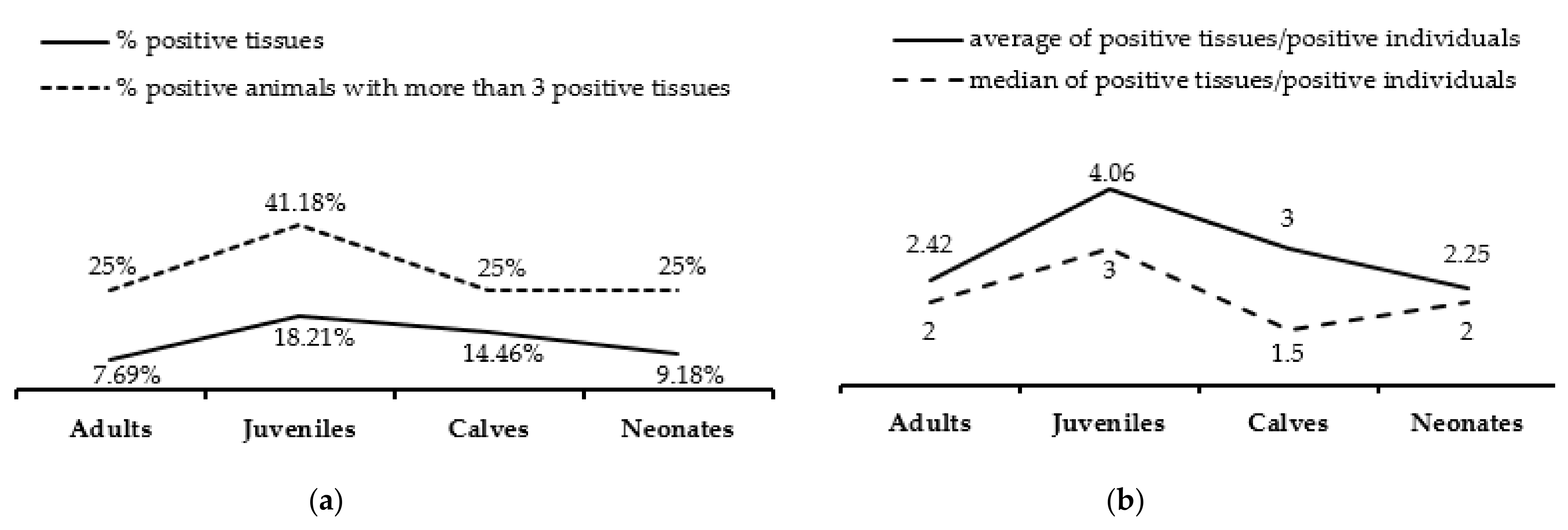
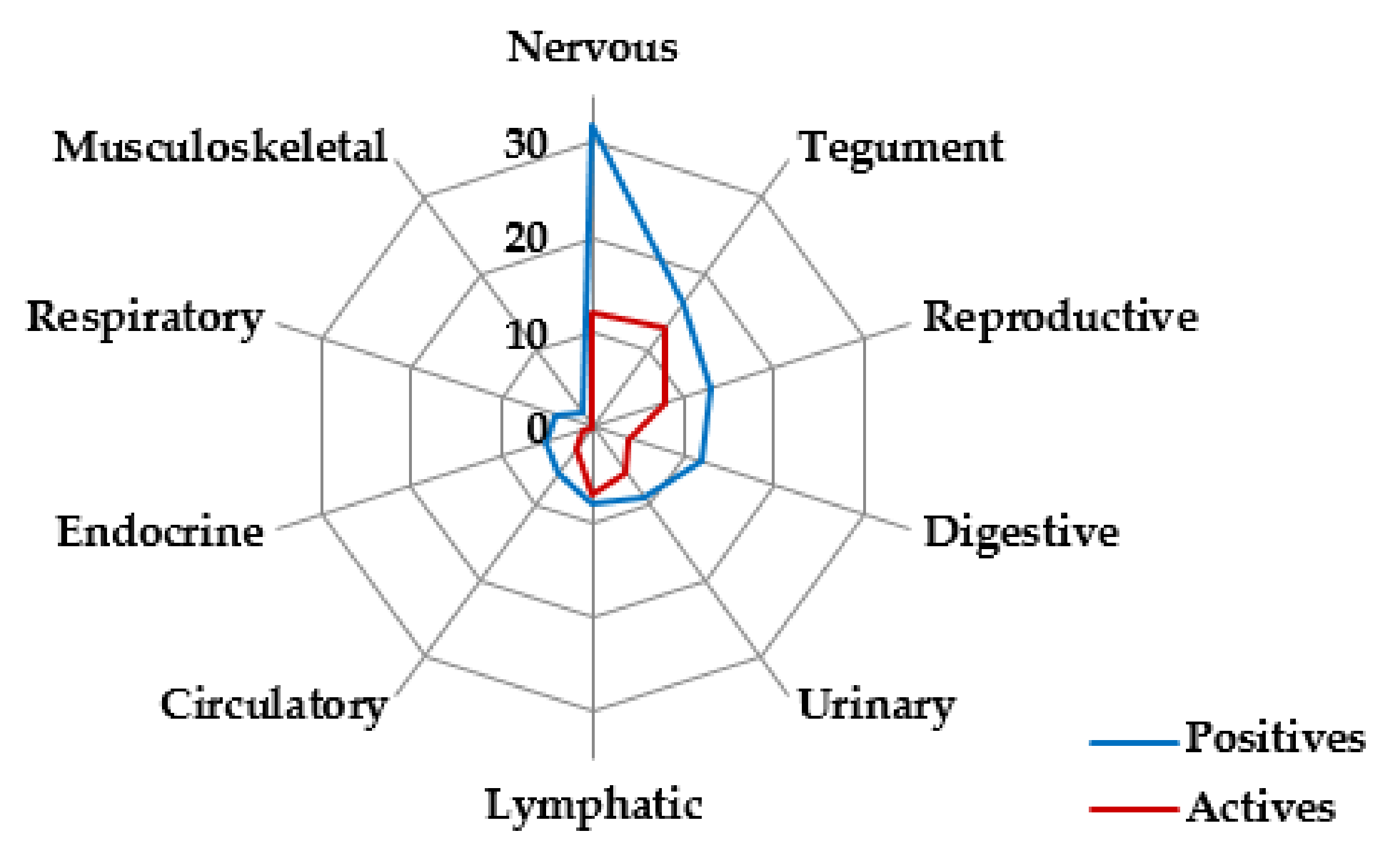
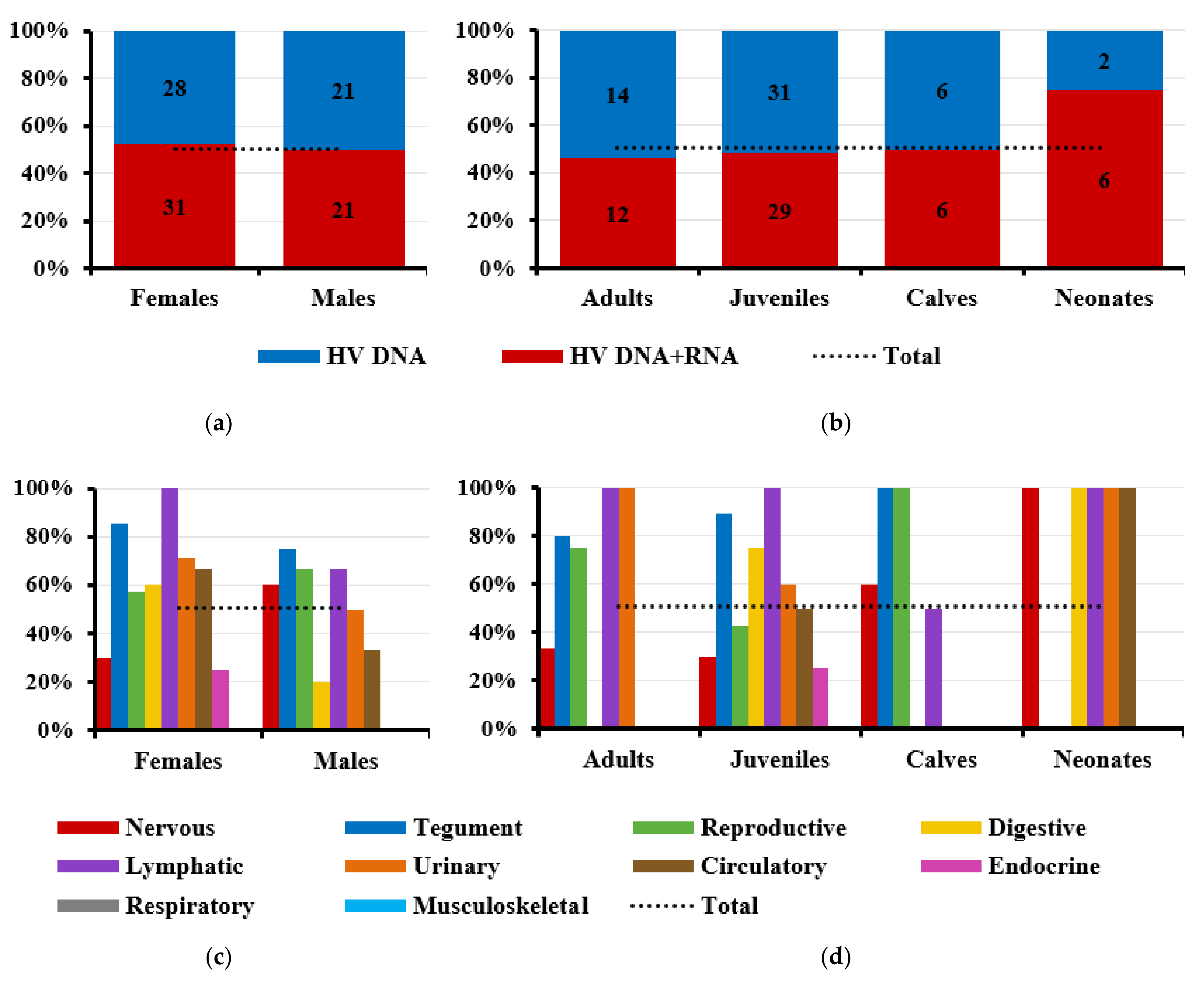
3.1. Herpesvirus Detection
3.2. Herpesvirus RNA Detection to Assess Active Replication
3.3. Phylogenetic Analysis
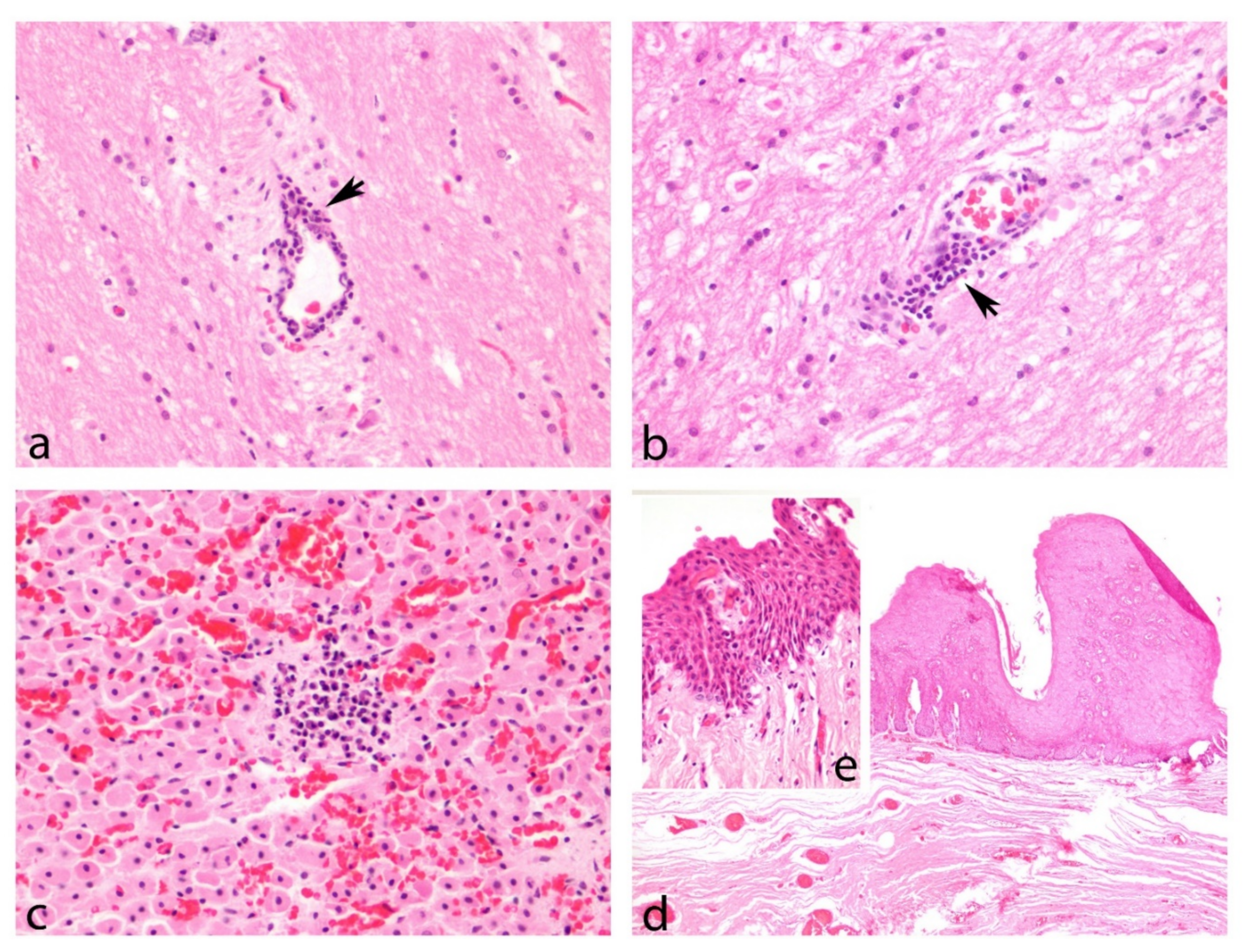
3.4. Gross Lesions and Histopathological Study
3.5. Herpesvirus and Morbillivirus Coinfection
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bossart, G.D. Marine mammals as sentinel species for oceans and human health. Vet. Pathol. 2011, 48, 676–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martineau, D.; Lagacé, A.; Béland, P.; Higgins, R.; Armstrong, D.; Shugart, L.R. Pathology of stranded beluga whales (Delphinapterus leucas) from the St. Lawrence Estuary, Québec, Canada. J. Comp. Pathol. 1988, 98, 287–310. [Google Scholar] [CrossRef]
- Barr, B.; Dunn, J.L.; Daniel, M.D.; Banford, A. Herpes-like viral dermatitis in a beluga whale (Delphinapterus leucas). J. Wildl. Dis. 1989, 25, 608–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, S.; Lindstedt, I.J.; McAliskey, M.M.; McConnell, S.A.; McCullough, S.J. Herpesviral encephalitis in a harbor porpoise (Phocoena phocoena). J. Zoo Wildl. Med. 1992, 23, 374–379. [Google Scholar]
- Van Bressem, M.F.; van Waerebeek, K.; Garcia-Godos, A.; Dekegel, D.; Pastoret, P.P. Herpes-like virus in Dusky dolphins, Lagenorhyncus obscurus, from coastal Peru. Mar. Mammal. Sci. 1994, 10, 354–359. [Google Scholar] [CrossRef]
- Blanchard, T.W.; Santiago, N.T.; Lipscomb, T.P.; Garber, R.L.; McFee, W.E.; Knowles, S. Two novel alphaherpesviruses associated with fatal disseminated infections in Atlantic bottlenose dolphins. J. Wildl. Dis. 2001, 37, 297–305. [Google Scholar] [CrossRef] [Green Version]
- Manire, C.A.; Smolarek, K.A.; Romero, C.H.; Kinsel, M.J.; Clauss, T.M.; Byrd, L. Proliferative dermatitis associated with a novel alphaherpesvirus in an Atlantic bottlenose dolphin (Tursiops Truncatus). J. Zoo Wildl. Med. 2006, 37, 174–181. [Google Scholar] [CrossRef]
- Saliki, J.T.; Cooper, E.J.; Rotstein, D.S.; Caseltine, S.L.; Pabst, D.A.; McLellan, W.A.; Govett, P.; Harms, C.; Smolarek, K.A.; Romero, C.H. A novel gammaherpesvirus associated with genital lesions in a Blainville’s beaked whale (Mesoplodon densirostris). J. Wildl. Dis. 2006, 42, 142–148. [Google Scholar] [CrossRef] [Green Version]
- Smolarek, K.A.; Manire, C.A.; Ewing, R.Y.; Saliki, J.T.; Townsend, F.I.; Ehlers, B.; Romero, C.H. Identification of novel alpha- and gammaherpesviruses from cutaneous and mucosal lesions of dolphins and whales. J. Virol. Methods 2006, 136, 261–266. [Google Scholar] [CrossRef]
- Esperón, F.; Fernández, A.; Sánchez-Vizcaíno, J.M. Herpes simplex-like infection in a bottlenose dolphin stranded in the Canary Islands. Dis. Aquat. Organ. 2008, 81, 73–76. [Google Scholar] [CrossRef] [Green Version]
- Van Elk, C.E.; van de Bildt, M.W.; de Jong, A.A.; Osterhaus, A.D.; Kuiken, T. Genital herpesvirus in bottlenose dolphins (Tursiops truncatus): Cultivation, epidemiology and associated pathology. J. Wildl. Dis. 2009, 45, 895–906. [Google Scholar] [CrossRef] [Green Version]
- Arbelo, M.; Sierra, E.; Esperón, F.; Watanabe, T.T.; Bellière, E.N.; Espinosa de los Monteros, A.; Fernández, A. Herpesvirus infection with severe lymphoid necrosis affecting a beaked whale stranded in the Canary Islands. Dis. Aquat. Organ. 2010, 89, 261–264. [Google Scholar] [CrossRef] [Green Version]
- Arbelo, M.; Bellière, E.N.; Sierra, E.; Sacchinni, S.; Esperón, F.; Andrada, M.; Rivero, M.; Diaz-Delgado, J.; Fernández, A. Herpes virus infection associated with interstitial nephritis in a beaked whale (Mesoplodon densirostris). BMC Vet. Res. 2012, 8, 243. [Google Scholar] [CrossRef] [Green Version]
- Soto, S.; González, B.; Willoughby, K.; Maley, M.; Olvera, A.; Kennedy, S.; Marco, A.; Domingo, M. Systemic herpesvirus and morbillivirus co-infection in a striped dolphin (Stenella coeruleoalba). J. Comp. Pathol. 2012, 146, 269–273. [Google Scholar] [CrossRef]
- Sierra, E.; Sánchez, S.; Saliki, J.T.; Blas-Machado, U.; Arbelo, M.; Zucca, D.; Fernández, A. Retrospective study of etiologic agents associated with nonsuppurative meningoencephalitis in stranded cetaceans in the Canary Islands. J. Clin. Microbiol. 2014, 52, 2390. [Google Scholar] [CrossRef] [Green Version]
- Bellehumeur, C.; Lair, S.; Romero, C.H.; Provost, C.; Nielsen, O.; Gagnon, C.A. Identification of a novel herpesvirus associated with a penile proliferative lesion in a beluga (Delphinapterus leucas). J. Wildl. Dis. 2015, 51, 244–249. [Google Scholar] [CrossRef] [Green Version]
- Melero, M.; Crespo-Picazo, J.L.; Rubio-Guerri, C.; García-Párraga, D.; Sánchez-Vizcaíno, J.M. First molecular determination of herpesvirus from two mysticete species stranded in the Mediterranean Sea. BMC Vet. Res. 2015, 11, 283. [Google Scholar] [CrossRef]
- Sierra, E.; Díaz-Delgado, J.; Arbelo, M.; Andrada, M.; Sacchini, S.; Fernández, A. Herpesvirus-associated genital lesions in a stranded striped dolphin (Stenella coeruleoalba) in the Canary Islands, Spain. J. Wildl. Dis. 2015, 51, 696–702. [Google Scholar] [CrossRef]
- Maness, H.T.; Nollens, H.H.; Jensen, E.D.; Goldstein, T.; LaMere, S.; Childress, A.; Sykes, J.; St Leger, J.; Lacave, G.; Latson, F.E.; et al. Phylogenetic analysis of marine mammal herpesviruses. Vet. Microbiol. 2011, 149, 23–29. [Google Scholar] [CrossRef]
- Bento, M.C.; Canha, R.; Eira, C.; Vingada, J.; Nicolau, L.; Ferreira, M.; Domingo, M.; Tavares, L.; Duarte, A. Herpesvirus infection in marine mammals: A retrospective molecular survey of stranded cetaceans in the Portuguese coastline. Infect. Genet. Evol. 2019, 67, 222–233. [Google Scholar] [CrossRef]
- Vargas-Castro, I.; Crespo-Picazo, J.L.; Rivera-Arroyo, B.; Sánchez, R.; Marco-Cabedo, V.; Jiménez-Martínez, M.Á.; Fayos, M.; Serdio, Á.; García-Párraga, D.; Sánchez-Vizcaíno, J.M. Alpha- and gammaherpesviruses in stranded striped dolphins (Stenella coeruleoalba) from Spain: First molecular detection of gammaherpesvirus infection in central nervous system of odontocetes. BMC Vet. Res. 2020, 16, 288. [Google Scholar] [CrossRef]
- Sacristán, C.; Esperón, F.; Ewbank, A.C.; Díaz-Delgado, J.; Ferreira-Machado, E.; Costa-Silva, S.; Sánchez-Sarmiento, A.M.; Groch, K.R.; Neves, E.; Pereira Dutra, G.H.; et al. Novel herpesviruses in riverine and marine cetaceans from South America. Acta Trop. 2019, 190, 220–227. [Google Scholar] [CrossRef]
- Van Elk, C.; van de Bildt, M.; van Run, P.; de Jong, A.; Getu, S.; Verjans, G.; Osterhaus, A.; Kuiken, T. Central nervous system disease and genital disease in harbor porpoises (Phocoena phocoena) are associated with different herpesviruses. Vet. Res. 2016, 47, 28. [Google Scholar] [CrossRef] [Green Version]
- Bellière, E.N.; Esperón, F.; Arbelo, M.; Muñoz, M.J.; Fernández, A.; Sánchez-Vizcaíno, J.M. Presence of herpesvirus in striped dolphins stranded during the cetacean morbillivirus epizootic along the Mediterranean Spanish coast in 2007. Arch. Virol. 2010, 155, 1307–1311. [Google Scholar] [CrossRef]
- Speck, S.H.; Ganem, D. Viral latency and its regulation: Lessons from the gamma-herpesviruses. Cell Host Microbe 2010, 8, 100–115. [Google Scholar] [CrossRef] [Green Version]
- Penkert, R.R.; Kalejta, R.F. Tegument protein control of latent herpesvirus establishment and animation. Herpesviridae 2011, 2, 3. [Google Scholar] [CrossRef] [Green Version]
- Glaser, R.; Kiecolt-Glaser, J.K. Stress-associated immune modulation and its implications for reactivation of latent herpesviruses. In Herpesvirus Infections; Glaser, R., Jones, J., Eds.; Springer: New York, NY, USA, 1994; pp. 245–270. [Google Scholar]
- Davison, A.J. Herpesvirus systematics. Vet. Microbiol. 2010, 143, 52–69. [Google Scholar] [CrossRef] [Green Version]
- Venn-Watson, S.; Benham, C.; Gulland, F.M.; Smith, C.R.; St Leger, J.; Yochem, P.; Nollens, H.; Blas-Machado, U.; Saliki, J.; Colegrove, K.; et al. Clinical relevance of novel Otarine herpesvirus-3 in California sea lions (Zalophus californianus): Lymphoma, esophageal ulcers, and strandings. Vet. Res. 2012, 43, 85. [Google Scholar] [CrossRef] [Green Version]
- Felipe-Jiménez, I.; Fernández, A.; Andrada, M.; Arbelo, M.; Segura-Göthlin, S.; Colom-Rivero, A.; Sierra, E. Contribution to Herpesvirus Surveillance in Beaked Whales Stranded in the Canary Islands. Animals 2021, 11, 1923. [Google Scholar] [CrossRef]
- Rehtanz, M.; Bossart, G.D.; Fair, P.A.; Reif, J.S.; Ghim, S.-J.; Jenson, A.B. Papillomaviruses and herpesviruses: Who is who in genital tumor development of free-ranging Atlantic bottlenose dolphins (Tursiops truncatus)? Vet. Microbiol. 2012, 160, 297–304. [Google Scholar] [CrossRef]
- Seade, G.C.C.; Cerqueira, V.D.; Sierra, E.; Chaves, J.F.; Moura, M.A.O.; Montão, D.P.; Riet-Correa, G.; Oliveira, C.A.; Siciliano, S.; Emin-Lima, R.; et al. Herpesviral infection in a Guiana dolphin (Sotalia guianensis) from the northern coast of Brazil. J. Vet. Diagn. Investig. 2017, 29, 877–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bigal, E.; Morick, D.; Scheinin, A.P.; Salant, H.; Berkowitz, A.; King, R.; Levy, Y.; Melero, M.; Sánchez-Vizcaíno, J.M.; Goffman, O.; et al. Detection of Toxoplasma gondii in three common bottlenose dolphins (Tursiops truncatus); A first description from the Eastern Mediterranean Sea. Vet. Parasitol. 2018, 258, 74–78. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, K.; Nishida, S.; Sone, E.; Tajima, Y.; Makara, M.; Yoshioka, M.; Nakamura, M.; Yamada, T.K.; Koike, H. Molecular Identification of Novel Alpha- and Gammaherpesviruses from Cetaceans Stranded on Japanese Coasts. Zool. Sci. 2011, 28, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Babiuk, L.A.; Lawman, M.J.P.; Griebel, P. Immunosuppression by bovine herpesvirus 1 and other selected herpesviruses. In Virus-Induced Immunosuppression; Specter, S., Bendinelli, M., Friedman, H., Eds.; Springer: Boston, MA, USA, 1989; pp. 141–171. [Google Scholar]
- Hannant, D.; O’Neill, T.; Ostlund, E.N.; Kydd, J.H.; Hopkin, P.J.; Mumford, J.A. Equid herpesvirus-induced immunosuppression is associated with lymphoid cells and not soluble circulating factors. Viral Immunol. 1999, 12, 313–321. [Google Scholar] [CrossRef]
- Islam, A.F.; Wong, C.W.; Walkden-Brown, S.W.; Colditz, I.G.; Arzey, K.E.; Groves, P.J. Immunosuppressive effects of Marek’s disease virus (MDV) and herpesvirus of turkeys (HVT) in broiler chickens and the protective effect of HVT vaccination against MDV challenge. Avian Pathol. 2002, 31, 449–461. [Google Scholar] [CrossRef]
- Flamand, L.; Gosselin, J.; Stefanescu, I.; Ablashi, D.; Menezes, J. Immunosuppressive effect of human herpesvirus 6 on T-cell functions: Suppression of interleukin-2 synthesis and cell proliferation. Blood 1995, 85, 1263–1271. [Google Scholar] [CrossRef]
- Dagna, L.; Pritchett, J.C.; Lusso, P. Immunomodulation and immunosuppression by human herpesvirus 6A and 6B. Future Virol. 2013, 8, 273–287. [Google Scholar] [CrossRef] [Green Version]
- Pellet, P.E.; Roizman, B. Herpesviridae. In Fields Virology, 6th ed.; Knipe, D.M., Howley, P.M., Eds.; Wolters Kluwer Health/Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; pp. 1802–1822. [Google Scholar]
- Mikaelian, I.; Tremblay, M.P.; Montpetit, C.; Tessaro, S.V.; Cho, H.J.; House, C.; Measures, L.; Martineau, D. Seroprevalence of selected viral infections in a population of beluga whales (Delphinapterus leucas) in Canada. Vet. Rec. 1999, 144, 50–51. [Google Scholar] [CrossRef]
- Domingo, M.; Visa, J.; Pumarola, M.; Marco, A.J.; Ferrer, L.; Rabanal, R.; Kennedy, S. Pathologic and immunocytochemical studies of morbillivirus infection in striped dolphins (Stenella coeruleoalba). Vet. Pathol. 1992, 29, 1–10. [Google Scholar] [CrossRef]
- Heaney, J.; Barrett, T.; Cosby, S.L. Inhibition of in vitro leukocyte proliferation by morbilliviruses. J. Virol. 2002, 76, 3579–3584. [Google Scholar] [CrossRef] [Green Version]
- Calzada, N.; Aguilar, A.; Grau, E.; Lockyer, C. Patterns of growth and physical maturity in the western Mediterranean striped dolphin, Stenella coeruleoalba (Cetacea: Odontoceti). Can. J. Zool. 1997, 75, 632–637. [Google Scholar] [CrossRef]
- Meissner, A.M.; MacLeod, C.D.; Richard, P.; Ridoux, V.; Pierce, G. Feeding ecology of striped dolphins, Stenella coeruleoalba, in the north-western Mediterranean Sea based on stable isotope analyses. J. Marine Biol. Assoc. UK 2012, 92, 1677–1687. [Google Scholar] [CrossRef] [Green Version]
- Chen, I.; Watson, A.; Chou, L.-S. Insights from life history traits of Risso’s dolphins (Grampus griseus) in Taiwanese waters: Shorter body length characterizes northwest Pacific population. Mar. Mammal. Sci. 2011, 27, E43–E64. [Google Scholar] [CrossRef]
- McFee, W.E.; Schwacke, J.H.; Stolen, M.K.; Mullin, K.D.; Schwacke, L.H. Investigation of growth phases for bottlenose dolphins using a Bayesian modeling approach. Mar. Mammal. Sci. 2010, 26, 67–85. [Google Scholar] [CrossRef] [Green Version]
- Read, A.J.; Wells, R.S.; Hohn, A.A.; Scott, M.D. Patterns of growth in wild bottlenose dolphins, Tursiops truncatus. J. Zool. 1993, 231, 107–123. [Google Scholar] [CrossRef]
- American Veterinary Medical Assoaciation. Report of the AVMA panel on euthanasia. J. Am. Vet. Med. Assoc. 1993, 202, 229–249. [Google Scholar]
- Close, B.; Banister, K.; Baumans, V.; Bernoth, E.M.; Bromage, N.; Bunyan, J.; Erhardt, W.; Flecknell, P.; Gregory, N.; Hackbarth, H.; et al. Recommendations for euthanasia of experimental animals: Part 1. DGXI of the European Commission. Lab. Anim. 1996, 30, 293–316. [Google Scholar] [CrossRef]
- Kuiken, T.; Hartmann, M.G. Cetacean Dissection techniques and tissue sampling: ECS newsletter special issue. In Proceedings of the Workshop, Leiden, The Netherlands, 13–14 September 1991; pp. 1–39. [Google Scholar]
- Geraci, J.; Lounsbury, V. Marine Mammals Ashore: A Field Guide for Strandings; National Aquarium in Baltimore: Baltimore, MA, USA, 2005. [Google Scholar]
- Van Devanter, D.R.; Warrener, P.; Bennett, L.; Schultz, E.R.; Coulter, S.; Garber, R.L.; Rose, T.M. Detection and analysis of diverse herpesviral species by consensus primer PCR. J. Clin. Microbiol. 1996, 34, 1666–1671. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.D.; Plewniak, F.; Poch, O. A comprehensive comparison of multiple sequence alignment programs. Nucleic Acids Res. 1999, 27, 2682–2690. [Google Scholar] [CrossRef]
- Ogden, T.H.; Rosenberg, M.S. Multiple Sequence Alignment Accuracy and Phylogenetic Inference. Syst. Biol. 2006, 55, 314–328. [Google Scholar] [CrossRef] [Green Version]
- Melero, M.; García-Párraga, D.; Corpa, J.M.; Ortega, J.; Rubio-Guerri, C.; Crespo, J.L.; Rivera-Arroyo, B.; Sánchez-Vizcaíno, J.M. First molecular detection and characterization of herpesvirus and poxvirus in a Pacific walrus (Odobenus rosmarus divergens). BMC Vet. Res. 2014, 10, 968. [Google Scholar] [CrossRef] [Green Version]
- Gröne, A.; Weckmann, M.T.; Capen, C.C.; Rosol, T.J. Canine glyceraldehyde-3-phosphate dehydrogenase complementary DNA: Polymerase chain reaction amplification, cloning, partial sequence analysis, and use as loading control in ribonuclease protection assays. Am. J. Vet. Res. 1996, 57, 254–257. [Google Scholar]
- Rubio-Guerri, C.; Melero, M.; Rivera-Arroyo, B.; Bellière, E.N.; Crespo, J.L.; García-Párraga, D.; Esperón, F.; Sánchez-Vizcaíno, J.M. Simultaneous diagnosis of Cetacean morbillivirus infection in dolphins stranded in the Spanish Mediterranean sea in 2011 using a novel Universal Probe Library (UPL) RT-PCR assay. Vet. Microbiol. 2013, 165, 109–114. [Google Scholar] [CrossRef]
- Rubio-Guerri, C.; Melero, M.; Esperón, F.; Bellière, E.N.; Arbelo, M.; Crespo, J.L.; Sierra, E.; García-Párraga, D.; Sánchez-Vizcaíno, J.M. Unusual striped dolphin mass mortality episode related to cetacean morbillivirus in the Spanish Mediterranean sea. BMC Vet. Res. 2013, 9, 106. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, T.; Gulland, F.M.D.; Braun, R.C.; Antonelis, G.A.; Kashinsky, L.; Rowles, T.K.; Mazet, J.A.; Dalton, L.M.; Aldridge, B.M.; Stott, J.L. Molecular identification of a novel gamma herpesvirus in the endangered Hawaiian monk seal (Monachus schauinslandi). Mar. Mammal Sci. 2006, 22, 465–471. [Google Scholar] [CrossRef]
- Zarnke, R.L.; Harder, T.C.; Vos, H.W.; ver Hoef, J.M.; Osterhaus, A.D. Serologic survey for phocid herpesvirus-1 and -2 in marine mammals from Alaska and Russia. J. Wildl. Dis. 1997, 33, 459–465. [Google Scholar] [CrossRef] [Green Version]
- Buckles, E.L.; Lowenstine, L.J.; DeLong, R.L.; Melin, S.R.; Vittore, R.K.; Wong, H.N.; Ross, G.L.; St Leger, J.A.; Greig, D.J.; Duerr, R.S.; et al. Age-prevalence of Otarine Herpesvirus-1, a tumor-associated virus, and possibility of its sexual transmission in California sea lions. Vet. Microbiol. 2007, 120, 1–8. [Google Scholar] [CrossRef]
- Cabello, J.; Esperón, F.; Napolitano, C.; Hidalgo, E.; Dávila, J.A.; Millán, J. Molecular identification of a novel gammaherpesvirus in the endangered Darwin’s fox (Lycalopex fulvipes). J. Gen. Virol. 2013, 94, 2745–2749. [Google Scholar] [CrossRef]
- Van Bressem, M.-F.; Duignan, P.J.; Banyard, A.; Barbieri, M.; Colegrove, K.M.; de Guise, S.; di Guardo, G.; Dobson, A.; Domingo, M.; Fauquier, D.; et al. Cetacean morbillivirus: Current knowledge and future directions. Viruses 2014, 6, 5145–5181. [Google Scholar] [CrossRef]
- Hierweger, M.M.; Boujon, C.L.; Kauer, R.V.; Meylan, M.; Seuberlich, T.; Oevermann, A. Cerebral Ovine Herpesvirus 2 Infection of Cattle Is Associated with a Variable Neuropathological Phenotype. Vet. Pathol. 2020, 58, 384–395. [Google Scholar] [CrossRef] [PubMed]
- Black, W.; Troyer, R.M.; Coutu, J.; Wong, K.; Wolff, P.; Gilbert, M.; Yuan, J.; Wise, A.G.; Wang, S.; Xu, D.; et al. Identification of gammaherpesvirus infection in free-ranging black bears (Ursus americanus). Virus Res. 2019, 259, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Sierra, E.; Fernández, A.; Felipe-Jiménez, I.; Zucca, D.; Díaz-Delgado, J.; Puig-Lozano, R.; Câmara, N.; Consoli, F.; Díaz-Santana, P.; Suárez-Santana, C.; et al. Histopathological Differential Diagnosis of Meningoencephalitis in Cetaceans: Morbillivirus, Herpesvirus, Toxoplasma gondii, Brucella sp., and Nasitrema sp. Front. Vet. Sci. 2020, 7, 650. [Google Scholar] [CrossRef] [PubMed]
- Van Beurden, S.J.; IJsseldijk, L.L.; Ordonez, S.R.; Förster, C.; de Vrieze, G.; Gröne, A.; Verheije, M.H.; Kik, M. Identification of a novel gammaherpesvirus associated with (muco)cutaneous lesions in harbour porpoises (Phocoena phocoena). Arch. Virol. 2015, 160, 3115–3120. [Google Scholar] [CrossRef]
- Burek-Huntington, K.A.; Dushane, J.L.; Goertz, C.E.; Measures, L.N.; Romero, C.H.; Raverty, S.A. Morbidity and mortality in stranded Cook Inlet beluga whales Delphinapterus leucas. Dis. Aquat. Organ. 2015, 114, 45–60. [Google Scholar] [CrossRef]
- Hart, L.B.; Rotstein, D.S.; Wells, R.S.; Allen, J.; Barleycorn, A.; Balmer, B.C.; Lane, S.M.; Speakman, T.; Zolman, E.S.; Stolen, M.; et al. Skin lesions on common bottlenose dolphins (Tursiops truncatus) from three sites in the Northwest Atlantic, USA. PLoS ONE 2012, 7, e33081. [Google Scholar] [CrossRef] [Green Version]
- Melero, M.; Rubio-Guerri, C.; Crespo, J.L.; Arbelo, M.; Vela, A.I.; García-Párraga, D.; Sierra, E.; Domínguez, L.; Sánchez-Vizcaíno, J.M. First case of erysipelas in a free-ranging bottlenose dolphin (Tursiops truncatus) stranded in the Mediterranean Sea. Dis. Aquat. Organ. 2011, 97, 167–170. [Google Scholar] [CrossRef] [Green Version]
- Mauldin, E.A.; Peters-Kennedy, J. Chapter 6—Integumentary system. In Jubb Kennedy and Palmer’s Pathology of Domestic Animals; Elsevier: New York, NY, USA, 2016; Volume 1, pp. 509–736. [Google Scholar] [CrossRef]
- Caswell, J.L.; Williams, K.J. Chapter 5—Respiratory system. In Jubb Kennedy and Palmer’s Pathology of Domestic Animals, 6th ed.; Maxie, M.G., Ed.; Elsevier: Saint louis, MO, USA, 2016; Volume 2, pp. 465–591.e464. [Google Scholar]
- Cocumelli, C.; Fichi, G.; Marsili, L.; Senese, M.; Cardeti, G.; Cersini, A.; Ricci, E.; Garibaldi, F.; Scholl, F.; di Guardo, G.; et al. Cetacean Poxvirus in Two Striped Dolphins (Stenella coeruleoalba) Stranded on the Tyrrhenian Coast of Italy: Histopathological, Ultrastructural, Biomolecular, and Ecotoxicological Findings. Front. Vet. Sci. 2018, 5, 219. [Google Scholar] [CrossRef]
- Maxie, M. Chapter 5—Male genital system. In Jubb, Kennedy & Palmer’s Pathology of Domestic Animals, 5th ed.; Maxie, M.G., Ed.; Elsevier: Philadelphia, PA, USA, 2007; p. 565. [Google Scholar]
- Van Bressem, M.F.; Raga, J.A.; di Guardo, G.; Jepson, P.D.; Duignan, P.J.; Siebert, U.; Barrett, T.; Santos, M.C.; Moreno, I.B.; Siciliano, S.; et al. Emerging infectious diseases in cetaceans worldwide and the possible role of environmental stressors. Dis. Aquat. Organ. 2009, 86, 143–157. [Google Scholar] [CrossRef]
- Cruz, D.; Rodríquez, M.; Kouri, V.; Soto, Y.; Zamora, L.; Rodríguez, D.; Barrera, M.; Rehtanz, M. Concurrent papillomavirus- and herpesvirus-infection in Atlantic bottlenose dolphins (Tursiops truncatus) inhabiting the Cuban coast. Mar. Mammal Sci. 2014, 30, 1564–1572. [Google Scholar] [CrossRef]
- Lecis, R.; Tocchetti, M.; Rotta, A.; Naitana, S.; Ganges, L.; Pittau, M.; Alberti, A. First gammaherpesvirus detection in a free-living Mediterranean bottlenose dolphin. J. Zoo Wildl. Med. 2014, 45, 922–925. [Google Scholar] [CrossRef]
- Ewing, R.Y.; Rotstein, D.S.; McLellan, W.A.; Costidis, A.M.; Lovewell, G.; Schaefer, A.M.; Romero, C.H.; Bossart, G.D. Macroscopic and Histopathologic Findings from a Mass Stranding of Rough-Toothed Dolphins (Steno bredanensis) in 2005 on Marathon Key, Florida, USA. Front. Vet. Sci. 2020, 7, 572. [Google Scholar] [CrossRef]
- Uzal, F.A.; Plattner, B.L.; Hostetter, J.M. Chapter 1—Alimentary system. In Jubb, Kennedy and Palmer’s Pathology of Domestic Animals, 6th ed.; Maxie, M.G., Ed.; Elsevier: Saint louis, MO, USA, 2016; pp. 1–257.e252. [Google Scholar]
- Gottschling, M.; Bravo, I.G.; Schulz, E.; Bracho, M.A.; Deaville, R.; Jepson, P.D.; Bressem, M.-F.V.; Stockfleth, E.; Nindl, I. Modular organizations of novel cetacean papillomaviruses. Mol. Phylogenet. Evol. 2011, 59, 34–42. [Google Scholar] [CrossRef]
- Borst, G.H.; Walvoort, H.C.; Reijnders, P.J.; van der Kamp, J.S.; Osterhaus, A.D. An outbreak of a herpesvirus infection in harbor seals (Phoca vitulina). J. Wildl. Dis. 1986, 22, 1–6. [Google Scholar] [CrossRef]
- Osterhaus, A.D.; Yang, H.; Spijkers, H.E.; Groen, J.; Teppema, J.S.; van Steenis, G. The isolation and partial characterization of a highly pathogenic herpesvirus from the harbor seal (Phoca vitulina). Arch. Virol. 1985, 86, 239–251. [Google Scholar] [CrossRef] [Green Version]
- Jaber, J.R.; Pérez, J.; Arbelo, M.; Andrada, M.; Hidalgo, M.; Gómez-Villamandos, J.C.; van den Ingh, T.; Fernández, A. Hepatic Lesions in Cetaceans Stranded in the Canary Islands. Vet. Pathol. 2004, 41, 147–153. [Google Scholar] [CrossRef]
- Cianciolo, R.E.; Mohr, F.C. Chapter 4—Urinary system. In Jubb, Kennedy and Palmer’s Pathology of Domestic Animals, 6th ed.; Maxie, M.G., Ed.; Elsevier: Saint louis, MO, USA, 2016; Volume 2, pp. 376–464.e371. [Google Scholar]
- Roperto, F.; Pratelli, A.; Guarino, G.; Ambrosio, V.; Tempesta, M.; Galati, P.; Iovane, G.; Buonavoglia, C. Natural Caprine Herpesvirus 1 (CpHV-1) Infection in Kids. J. Comp. Pathol. 2000, 122, 298–302. [Google Scholar] [CrossRef]
- Groch, K.R.; Santos-Neto, E.B.; Díaz-Delgado, J.; Ikeda, J.M.P.; Carvalho, R.R.; Oliveira, R.B.; Guari, E.B.; Bisi, T.L.; Azevedo, A.F.; Lailson-Brito, J.; et al. Guiana Dolphin Unusual Mortality Event and Link to Cetacean Morbillivirus, Brazil. Emerg. Infect. Dis. 2018, 24, 1349–1354. [Google Scholar] [CrossRef] [Green Version]
- Sierra, E.A.M.; Herraez, P.; Esperon, F.; Sanchez-Vizcaino, J.M.; y Fernandez, A. Adrenalitis associated with a herpesvirus infection in a stranded bottlenose dolphin in Canary Islands. In Proceedings of the XX Reunión de la SEAPV (Sociedad Española de Anatomía Patológica Veterinaria), Breña Baja, Spain, 18–20 June 2008. [Google Scholar]
- Rosol, T.J.; Gröne, A. Chapter 3—Endocrine glands. In Jubb, Kennedy and Palmer’s Pathology of Domestic Animals, 6th ed.; Maxie, M.G., Ed.; Elsevier: Saint louis, MO, USA, 2016; Volume 3, pp. 269–357.e261. [Google Scholar]
- Tischer, B.K.; Osterrieder, N. Herpesviruses—A zoonotic threat? Vet. Microbiol. 2010, 140, 266–270. [Google Scholar] [CrossRef] [Green Version]
- Weidner-Glunde, M.; Kruminis-Kaszkiel, E.; Savanagouder, M. Herpesviral Latency-Common Themes. Pathogens 2020, 9, 125. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
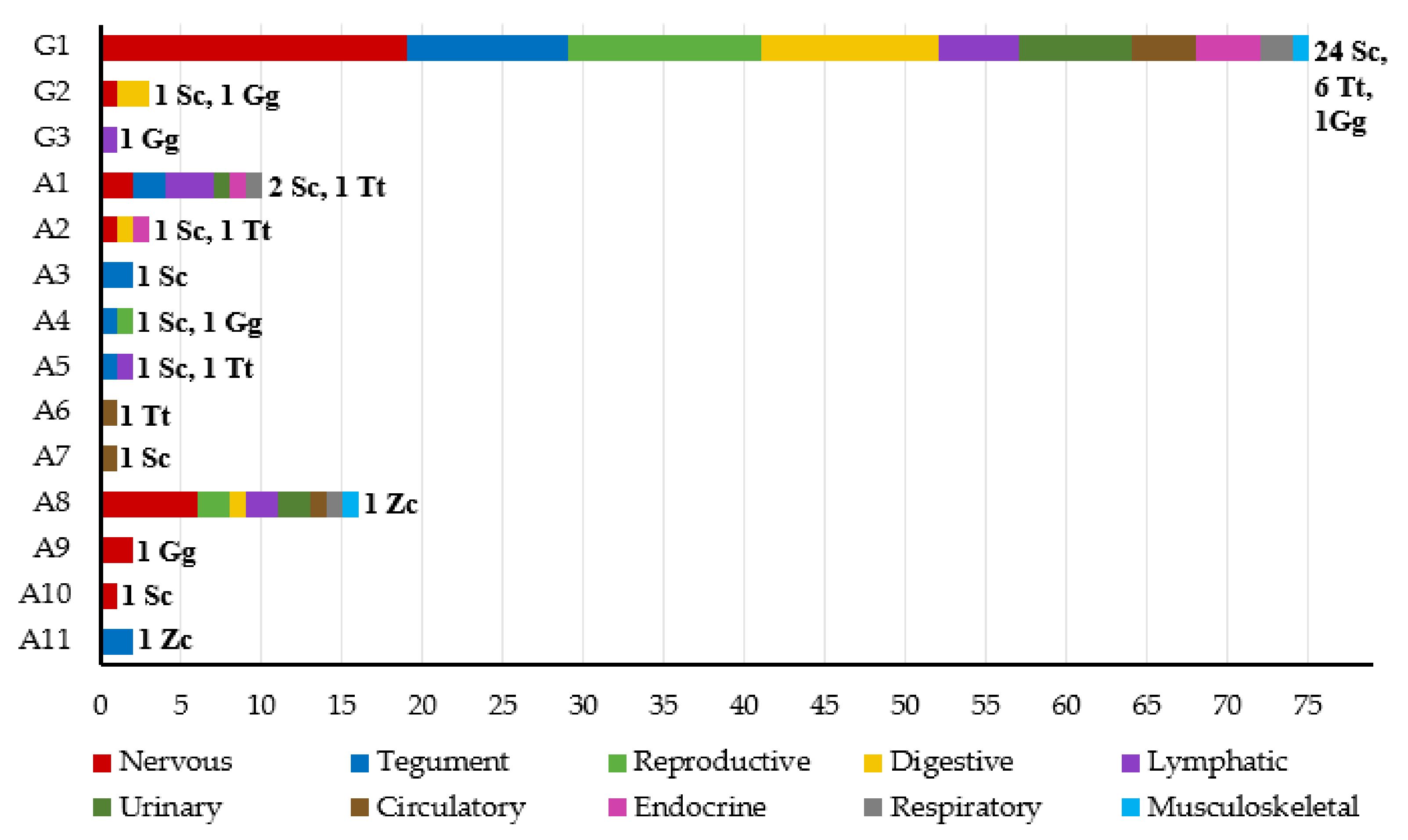{kind=link}
{kind=link}
{kind=link}
| Striped Dolphin | Bottlenose Dolphin | Risso’s Dolphin | Cuvier’s Beaked Whale | Total | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 35 Sc | 8 Tt | 3 Gg | 1 Zc | 47 Individuals | ||||||||||
| Tissue | 694 Tissues | 149 Tissues | 100 Tissues | 23 Tissues | 966 Tissues | |||||||||
| Reproductive system (39.47%) | Mammary gland | 1 | 2 (1G) | - | - | 3 (1G) | ||||||||
| 1G1 | ||||||||||||||
| Uterus | - | - | - | 1 (1A) | 1 (1A) | |||||||||
| 1A8 | ||||||||||||||
| Ovary | 3 (1G) | - | 1 (1G) | - | 4 (2G) | |||||||||
| 1G1 | 1G1 | |||||||||||||
| Testicle | 4 | 1 | - | - | 5 | |||||||||
| Penis | 7 (3G) | 3 (1G) | 2 (1A) | - | 12 (1A, 4G) | |||||||||
| 3G1 | 1G1 | 1A4 | ||||||||||||
| External genital mucosa | 7 (2G) | 2 (2G) | 2 (1G) | 1 (1A) | 12(1A, 5G) | |||||||||
| 2G1 | 2G1 | 1G1 | 1A8 | |||||||||||
| External genital secretion | - | - | 1 | - | 1 | |||||||||
| Nervous System (18.93%) | Optic nerve | 4 | - | - | - | 4 | ||||||||
| Meninges | 2 | 4 (1A) | 1 | - | 7 (1A) | |||||||||
| 1A2 | ||||||||||||||
| Cerebral cortex | 1 | - | 1 | 1 (1A) | 3 (1A) | |||||||||
| 1A8 | ||||||||||||||
| Brain (unspecified location) | 34 (6G) | 7 (2G) | 2 (1A) | - | 43 (1A, 8G) | |||||||||
| 5G1 | 1G2 | 2G1 | 1A9 | |||||||||||
| Lateral ventricle | 12 (1G) | 2 (1G) | 1 | 1 (1A) | 16 (1A, 2G) | |||||||||
| 1G1 | 1G1 | 1A8 | ||||||||||||
| Thalamus | 14 (2G) | 2 | 3 | 1 (1A) | 20 (1A, 2G) | |||||||||
| 2G1 | 1A8 | |||||||||||||
| Pons | 17 (2A, 1G) | 2 (1G) | 3 | 1 (1A) | 23 (3A, 2G) | |||||||||
| 1A1 | 1A10 | 1G1 | 1G1 | 1A8 | ||||||||||
| Cerebellum | 23 (1A, 3G) | 5 (1G) | 3 (1A) | 1 (1A) | 32 (3A, 4G) | |||||||||
| 1A1 | 3G1 | 1G1 | 1A9 | 1A8 | ||||||||||
| Spinal cord | 15 (2G) | 2 | 3 | 1 (1A) | 21 (1A, 2G) | |||||||||
| 2G1 | 1A8 | |||||||||||||
| Tegument (14.40%) | Epidermis | 62 (4A, 5G) | 14 (1A, 3G) | 6 | 1 (1A) | 83 (6A, 8G) | ||||||||
| 1A1 | 2A3 | 1A4 | 5G1 | 1A5 | 3G1 | 1A11 | ||||||||
| Hipodermis | 31 (1A, 2G) | 7 | 3 | 1 (1A) | 42 (2A, 2G) | |||||||||
| 1A1 | 2G1 | 1A11 | ||||||||||||
| Endocrine system (13.95%) | Thyroid | 9 (1G) | 1 | 2 | - | 12 (1G) | ||||||||
| 1G1 | ||||||||||||||
| Adrenal gland | 23 (2A, 3G) | 4 | 3 | 1 | 31 (2A, 3G) | |||||||||
| 1A1 | 1A2 | 3G1 | ||||||||||||
| Circulatory system (12.28%) | Myocardium | 13 (2G) | 2 | 3 (1G) | 1 (1A) | 19 (1A, 3G) | ||||||||
| 2G1 | 1G1 | 1A8 | ||||||||||||
| Heart valve | 2 | 2 | 1 | - | 5 | |||||||||
| Pericardial sac | 1 | - | - | - | 1 | |||||||||
| Pericardial fluid | 1 | 2 | - | - | 3 | |||||||||
| Blood | 12 (1A, 1G) | 7 (1A) | 3 | - | 22 (2A, 1G) | |||||||||
| 1A7 | 1G1 | 1A6 | ||||||||||||
| Serum | 4 | 1 | 2 | - | 7 | |||||||||
| Urinary apparatus (11.63%) | Kidney | 30 (1A) | 7 | 4 | 1 (1A) | 42 (2A) | ||||||||
| 1A1 | 1A8 | |||||||||||||
| Urinary bladder | 25 (5G) | 5 | 3 (1G) | 1 (1A) | 34 (1A, 6G) | |||||||||
| 5G1 | 1G1 | 1A8 | ||||||||||||
| Urine | 2 | 4 | 3 (1G) | 1 | 10 (1G) | |||||||||
| 1G1 | ||||||||||||||
| Digestive system (10.13%) | Oral cavity | Teeth and gingiva | 30(4G) | 7(1A) | 2 | - | 39 (1A, 4G) | |||||||
| 4G1 | 1A2 | |||||||||||||
| Tongue | 16 | - | 3 (1G) | - | 19 (1G) | |||||||||
| 1G1 | ||||||||||||||
| Oral swab | 3 | - | 2 | - | 5 | |||||||||
| Digestive tube | Esophagus | - | - | 2 (2G) | - | 2 (2G) | ||||||||
| 2G2 | ||||||||||||||
| Intestine | 7 | 1 | 1 | - | 9 | |||||||||
| Liver | 31 (2G) | 7 (2G) | 3 | 1 | 42 (4G) | |||||||||
| 2G1 | 2G1 | |||||||||||||
| Bile | 1 | - | - | - | 1 | |||||||||
| Pancreas | 24 (2G) | 3 | 3 | 1 (1A) | 31 (1A, 2G) | |||||||||
| 2G1 | 1A8 | |||||||||||||
| Respiratory system (6.78%) | Blowhole swab | 2 | - | 2 | - | 4 | ||||||||
| Lung | 39 (1A, 2G) | 8 | 5 | 1 (1A) | 53 (2A, 2G) | |||||||||
| 1A1 | 2G1 | 1A8 | ||||||||||||
| Lung exudate | 2 | - | - | - | 2 | |||||||||
| Lymphatic system (4.64%) | Spleen | 25 | 4 | 3 | 1 | 33 | ||||||||
| Thymus | 19 (1A) | 4 | 1 | - | 24 (1A) | |||||||||
| 1A5 | ||||||||||||||
| Pharyngeal tonsils | 21 (1A, 2G) | 2 | 4 (2G) | - | 27 (1A, 4G) | |||||||||
| 1A1 | 2G1 | 1G1 | 1G3 | |||||||||||
| Lymph nodes | Prescapular | 26 (1A, 1G) | 6 | 3 | 1 (1A) | 36 (2A, 1G) | ||||||||
| 1A1 | 1G1 | 1A8 | ||||||||||||
| Pulmonar | 32 | 6 | 3 | 1 (1A) | 42 (1A) | |||||||||
| 1A8 | ||||||||||||||
| Mesenteric | 23 (1G) | 5 (1A) | 3 | 1 | 32 (1A, 1G) | |||||||||
| 1G1 | 1A1 | |||||||||||||
| Musculoskeletal system (4.54%) | Epaxial muscle, thoracic region | 32 | 8 (1G) | 3 | 1 (1A) | 44 (1A, 1G) | ||||||||
| 1G1 | 1A8 | |||||||||||||
| Sense organs (0%) | Eye | 2 | - | 1 | - | 3 | ||||||||
| TOTAL | Positive tissues | 70 of 694 (16A 54G) 10.09% | 20 of 149 (5A 15G) 13.42% | 13 of 100 (3A 10G) 13.00% | 18 of 23 (18A) 78.26% | 121 of 966 (42A, 79G) 12.53% | ||||||||
| Positive individuals | 28 of 35 80.00% | 7 of 8 87.50% | 2 of 3 66.67% | 1 of 1 100% | 38 of 47 80.85% | |||||||||
| Stranding Date | Species | Sex | Age | Sample | HVSF | HV Seq | RNA | Concomitant Lesions | Possible Differential Diagnosis |
|---|---|---|---|---|---|---|---|---|---|
| 25.06.10 | Tt | M | A | Liver | GHV | G1 | - | Suppurative hepatitis | Bacterial |
| 14.07.10 | Tt | F | A | Urinary bladder | GHV | G1 | - | None | - |
| CNS | GHV | G1 | + | None | - | ||||
| Mammary gland | GHV | G1 | + | None | - | ||||
| Vagina | GHV | G1 | + | Lymphoplasmacytic and hiperplastic vaginitis | Cetacean papillomavirus | ||||
| Skin | GHV | G1 | + | Hyperplastic dermatitis | Cetacean poxvirus | ||||
| Skeletal muscle | GHV | G1 | - | None | - | ||||
| Lymph nodes | GHV | G1 | + | None | - | ||||
| Adrenal gland | GHV | G1 | + | None | - | ||||
| 14.08.10 | Sc | U | J | Skin | GHV | G1 | + | Hyperplastic dermatitis with parasitic granuloma (nematodes) | - |
| 12.03.11 | Sc | M | N | Myocardium | AHV | A7 | + | Degeneration | - |
| 13.03.11 | Sc | M | N | Lung | GHV | G1 | - | Chronic interstitial pneumonia | CeMV |
| 15.03.11 | Sc | F | J | Urinary bladder | GHV | G1 | - | Hemorrhagic cystitis | - |
| Skin | GHV | G1 | + | None | - | ||||
| Adrenal glands | GHV | G1 | + | Bilateral lymphoplasmacytic adrenalitis | - | ||||
| 16.03.11 | Sc | F | A | Urinary bladder | GHV | G1 | + | Lymphoplasmacytic cystitis with intracytoplasmic inclusion bodies | CeMV |
| 23.03.11 | Sc | F | A | CNS | GHV | G1 | + | Lymphoplasmacytic meningoencephalitis | CeMV, Brucella ceti, Toxoplasma gondii |
| 23.03.11 | Tt | M | J | CNS | AHV | A2 | + | Lymphoplasmacytic meningitis | CeMV, Brucella ceti |
| Skeletal muscle | GHV | G1 | - | None | - | ||||
| 25.03.11 | Sc | F | J | CNS | GHV | G1 | + | Lymphoplasmacytic meningoencephalitis | CeMV, Brucella ceti, Toxoplasma gondii |
| Skin | GHV | G1 | + | Degenerative and necrotizing dermatitis | - | ||||
| Adrenal gland | AHV | A2 | - | None | - | ||||
| 26.03.11 | Sc | F | J | CNS | GHV | G1 | + | Lymphoplasmacytic meningoencephalitis | CeMV, Brucella ceti, Toxoplasma gondii |
| Skin | GHV | G1 | - | Lymphoplasmacytic dermatitis | - | ||||
| 29.03.11 | Sc | M | C | CNS | GHV | G1 | - | Lymphoplasmacytic meningoencephalitis | - |
| 18.06.11 | Sc | M | J | Skin | AHV | A3 | + | Hyperplastic dermatitis | Cetacean poxvirus |
| 07.07.11 | Sc | M | J | CNS | GHV | G1 | + | Lymphoplasmacytic meningoencephalitis | CeMV, Brucella ceti, Toxoplasma gondii |
| Skin | GHV | G1 | + | Hyperplastic dermatitis | - | ||||
| 29.07.11 | Sc | F | A | Pancreas | GHV | G1 | - | None | - |
| Liver | GHV | G1 | - | Chronic cholangiohepatitis | Trematode | ||||
| 12.10.11 | Sc | M | J | Urinary bladder | GHV | G1 | + | Lymphoplasmacytic cystitis | - |
| Vaginal mucosa | GHV | G1 | + | Lymphoplasmacytic vaginitis | - | ||||
| 11.12.11 | Gg | M | A | Penis | AHV | A4 | - | Proliferative balanitis | Cetacean papillomavirus |
| Esophagus | GHV | G2 | - | Proliferative esophagitis | Cetacean papillomavirus | ||||
| 19.12.11 | Sc | F | C | CNS | GHV | G2 | - | Lymphoplasmacytic meningitis | CeMV, Brucella ceti |
| 26.01.12 | Sc | M | C | Kidney | AHV | A1 | - | Lymphoplasmacytic nephritis | - |
| CNS | AHV | A1 | + | None | - | ||||
| Lung | AHV | A1 | - | Interstitial bronchopneumonia with associated granulomatous bronchopneumonia | CeMV, Nematodes | ||||
| Lymph node | AHV | A1 | - | Granulomatous lymphadenitis | Parasitic migration | ||||
| 22.03.12 | Sc | F | J | CNS | GHV | G1 | - | Lymphoplasmacytic meningoencephalitis | CeMV, Brucella ceti, Toxoplasma gondii |
| Myocardium | GHV | G1 | - | None | - | ||||
| 15.05.12 | Zc | F | J | Kidney | AHV | A8 | - | Interstitial nephritis | - |
| CNS | AHV | A8 | + | None | - | ||||
| Lung | AHV | A8 | - | Fibrinonecrotic bronchopneumonia | Aspiration of fibrinonecrotic content | ||||
| Uterus | AHV | A8 | - | None | - | ||||
| Skin | AHV | A11 | - | None | - | ||||
| Lymph node | AHV | A8 | - | Lymphoid depletion | - | ||||
| Myocardium | AHV | A8 | - | None | - | ||||
| 07.09.12 | Sc | M | N | Liver | GHV | G1 | - | None | - |
| 20.10.12 | Sc | M | J | Myocardium | GHV | G1 | - | None | - |
| 13.02.13 | Sc | M | A | Skin | AHV | A4 | + | Hyperplastic dermatitis | Cetacean poxvirus |
| 29.03.13 | Gg | F | J | Urinary bladder | GHV | G1 | + | None | - |
| CNS | AHV | A9 | - | None | - | ||||
| Myocardium | GHV | G1 | - | None | - | ||||
| 12.05.13 | Sc | M | J | Penis | GHV | G1 | + | None | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vargas-Castro, I.; Melero, M.; Crespo-Picazo, J.L.; Jiménez, M.d.l.Á.; Sierra, E.; Rubio-Guerri, C.; Arbelo, M.; Fernández, A.; García-Párraga, D.; Sánchez-Vizcaíno, J.M. Systematic Determination of Herpesvirus in Free-Ranging Cetaceans Stranded in the Western Mediterranean: Tissue Tropism and Associated Lesions. Viruses 2021, 13, 2180. https://doi.org/10.3390/v13112180
Vargas-Castro I, Melero M, Crespo-Picazo JL, Jiménez MdlÁ, Sierra E, Rubio-Guerri C, Arbelo M, Fernández A, García-Párraga D, Sánchez-Vizcaíno JM. Systematic Determination of Herpesvirus in Free-Ranging Cetaceans Stranded in the Western Mediterranean: Tissue Tropism and Associated Lesions. Viruses. 2021; 13(11):2180. https://doi.org/10.3390/v13112180
Chicago/Turabian StyleVargas-Castro, Ignacio, Mar Melero, José Luis Crespo-Picazo, María de los Ángeles Jiménez, Eva Sierra, Consuelo Rubio-Guerri, Manuel Arbelo, Antonio Fernández, Daniel García-Párraga, and José Manuel Sánchez-Vizcaíno. 2021. "Systematic Determination of Herpesvirus in Free-Ranging Cetaceans Stranded in the Western Mediterranean: Tissue Tropism and Associated Lesions" Viruses 13, no. 11: 2180. https://doi.org/10.3390/v13112180